La fotosíntesis es la conversión de energía luminosa en energía química estable, siendo el adenosín trifosfato (ATP) la primera molécula en la que queda almacenada esa energía química. Con posterioridad, el ATP se usa para sintetizar moléculas orgánicas de mayor estabilidad. Además, se debe de tener en cuenta que la vida en nuestro planeta se mantiene fundamentalmente gracias a la fotosíntesis que realizan las algas, en el medio acuático, y las plantas, en el medio terrestre, que tienen la capacidad de sintetizar materia orgánica (imprescindible para la constitución de los seres vivos) partiendo de la luz y la materia inorgánica. De hecho, cada año los organismos fotosintetizadores fijan en forma de materia orgánica en torno a 100.000 millones de toneladas de carbono.
Los orgánulos citoplasmáticos encargados de la realización de la fotosíntesis son los cloroplastos, unas estructuras polimorfas y de color verde (esta coloración es debida a la presencia del pigmento clorofila) propias de las células vegetales. En el interior de estos orgánulos se halla una cámara que contiene un medio interno llamado estroma, que alberga diversos componentes, entre los que cabe destacar enzimas encargadas de la transformación del dióxido de carbono en materia orgánica y unos sáculos aplastados denominados tilacoides o lamelas, cuya membrana contiene pigmentos fotosintéticos. En términos medios, una célula foliar tiene entre cincuenta y sesenta cloroplastos en su interior.
Los organismos que tienen la capacidad de llevar a cabo la fotosíntesis son llamados fotoautótrofos (otra nomenclatura posible es la de autótrofos, pero se debe tener en cuenta que bajo esta denominación también se engloban aquellas bacterias que realizan la quimiosíntesis) y fijan el CO2 atmosférico. En la actualidad se diferencian dos tipos de procesos fotosintéticos, que son la fotosíntesis oxigénica y la fotosíntesis anoxigénica. La primera de las modalidades es la propia de las plantas superiores, las algas y las cianobacterias, donde el dador de electrones es el agua y, como consecuencia, se desprende oxígeno. Mientras que la segunda, también conocida con el nombre de fotosíntesis bacteriana, la realizan las bacterias purpúreas y verdes del azufre, en las que en dador de electrones es el sulfuro de hidrógeno, y consecuentemente, el elemento químico liberado no será oxígeno sino azufre, que puede ser acumulado en el interior de la bacteria, o en su defecto, expulsado al agua.
A comienzos del año 2009, se publicó un artículo en la revista Nature Geoscience en el que científicos norteamericanos daban a conocer el hallazgo de pequeños cristales de hematita (en Cratón de Pilbara, en el noroeste de Australia), un mineral de hierro que data de la época del eón Arcaico, demostrando la existencia de agua rica en oxígeno y consecuentemente, de organismos fotosintetizadores capaces de producirlo. Gracias al estudio realizado, se ha llegado a la conclusión de la existencia de fotosíntesis oxigénica y de la oxigenación de la atmósfera y de los océanos hace más de 3.460 millones de años, así como también se deduce la existencia de un número considerable de organismos capaces de llevar a cabo la fotosíntesis para oxigenar la masa de agua mencionada, aunque sólo fuese de manera ocasional.
martes, 9 de noviembre de 2010
EL CLOROPLASTO
De todas las células eucariotas, únicamente las fotosintéticas presentan cloroplastos, unos orgánulos que usan la energía solar para impulsar la formación de ATP y NADH, compuestos utilizados con posterioridad para el ensamblaje de azúcares y otros compuestos orgánicos. Al igual que las mitocondrias, cuentan con su propio ADN y posiblemente se hayan originado como bacterias simbióticas intracelulares.
 En las células meristemáticas se encuentran proplastos, que no tienen ni membrana interna, ni clorofila, ni ciertos enzimas requeridos para llevar a cabo la fotosíntesis. En angiospermas y gimnospermas el desarrollo de los cloroplastos es desencadenado por la luz, puesto que bajo iluminación se generan los enzimas en el interior del proplasto o se extraen del citosol, aparecen los pigmentos encargados de la absorción lumínica y se producen con gran rapidez las membranas, dando lugar a los grana y las lamelas del estroma.
En las células meristemáticas se encuentran proplastos, que no tienen ni membrana interna, ni clorofila, ni ciertos enzimas requeridos para llevar a cabo la fotosíntesis. En angiospermas y gimnospermas el desarrollo de los cloroplastos es desencadenado por la luz, puesto que bajo iluminación se generan los enzimas en el interior del proplasto o se extraen del citosol, aparecen los pigmentos encargados de la absorción lumínica y se producen con gran rapidez las membranas, dando lugar a los grana y las lamelas del estroma.
A pesar de que las semillas suelen germinar en el suelo sin luz, los cloroplastos son una clase de orgánulos que exclusivamente se desarrollan cuando el vástago queda expuesto a la luz. Si la semilla germina en ausencia de luz, los proplastos se diferencian en etioplastos, que albergan una agrupación tubular semicristalina de membrana llamada cuerpo prolamelar. En vez de clorofila, estos etioplastos tienen un pigmento de color verde-amarillento que constituye el precursor de la misma: es la denominada protoclorofila.
Después de estar por un pequeño intervalo de tiempo expuestos a la luz, los etioplastos se diferencian transformándose los cuerpos prolamelares en tilacoides y lamelas del estroma, y la protoclorofila, en clorofila. El mantenimiento de la estructura de los cloroplastos está directamente vinculada a la luz, de modo que si en algún momento éstos pasan a estar en penumbra continuada puede desencadenarse que los cloroplastos vuelvan a convertirse en etioplastos.
Además, los cloroplastos pueden convertirse en cromoplastos, como sucede en las hojas durante el otoño o a lo largo del proceso de maduración de los frutos (proceso reversible en determinadas ocasiones). Asimismo, los amiloplastos (contenedores de almidón) pueden transformarse en cloroplastos, hecho que explica el fenómeno por el cual las raíces adquieren tonos verdosos al estar en contacto con la luz solar.
 Se distinguen por ser unas estructuras polimorfas de color verde, siendo la coloración que presentan consecuencia directa de la presencia del pigmento clorofila en su interior. Además, presentan una envoltura formada por una doble membrana que carece de clorofila y colesterol: una membrana plastidial externa y una membrana plastidial interna.
Se distinguen por ser unas estructuras polimorfas de color verde, siendo la coloración que presentan consecuencia directa de la presencia del pigmento clorofila en su interior. Además, presentan una envoltura formada por una doble membrana que carece de clorofila y colesterol: una membrana plastidial externa y una membrana plastidial interna.
En las plantas superiores, la forma que con mayor frecuencia presentan los cloroplastos es la de disco lenticular, aunque también existen algunos de aspecto ovoidal o esférico. Con respecto a su número, se puede decir que en torno a cuarenta y cincuenta cloroplastos coexisten, de media, en una célula de una hoja; y existen unos 500.000 cloroplastos por milímetro cuadrado de superficie foliar. No sucede lo mismo entre las algas, pues los cloroplastos de éstas no se encuentran tan determinados ni en número ni en forma. Por ejemplo, en el alga Spirogyra únicamente existen dos cloroplastos con forma de cinta en espiral, y en el alga Chlamydomonas, sólo hay uno de grandes dimensiones.
En el interior y delimitado por una membrana plastidial interna, se ubica una cámara que alberga un medio interno con un elevado número de componentes (ADN plastidial, circular y de doble hélice, plastorribosomas, enzimas e inclusiones de granos de almidón y las inclusiones lipídicas); es lo que se conoce por el nombre de estroma. Inmerso en el se encuentran una gran cantidad de sáculos denominados tilacoides, que contienen pigmentos fotosintéticos en su membrana tilacoidal (cuya cavidad interior se llama lumen o espacio tilacoidal). Los tilacoides pueden encontrarse repartidos por todo el estroma (tilacoides del estroma), o bien, pueden ser pequeños, tener forma discoidal y encontrarse apilados originando unos montones, denominados grana (tilacoides de grana). Es en la membrana de los grana donde se ubican los sistemas enzimáticos encargados de captar la energía luminosa, llevar a cabo el transporte de electrones y sintetizar ATP.

A pesar de que las semillas suelen germinar en el suelo sin luz, los cloroplastos son una clase de orgánulos que exclusivamente se desarrollan cuando el vástago queda expuesto a la luz. Si la semilla germina en ausencia de luz, los proplastos se diferencian en etioplastos, que albergan una agrupación tubular semicristalina de membrana llamada cuerpo prolamelar. En vez de clorofila, estos etioplastos tienen un pigmento de color verde-amarillento que constituye el precursor de la misma: es la denominada protoclorofila.
Después de estar por un pequeño intervalo de tiempo expuestos a la luz, los etioplastos se diferencian transformándose los cuerpos prolamelares en tilacoides y lamelas del estroma, y la protoclorofila, en clorofila. El mantenimiento de la estructura de los cloroplastos está directamente vinculada a la luz, de modo que si en algún momento éstos pasan a estar en penumbra continuada puede desencadenarse que los cloroplastos vuelvan a convertirse en etioplastos.
Además, los cloroplastos pueden convertirse en cromoplastos, como sucede en las hojas durante el otoño o a lo largo del proceso de maduración de los frutos (proceso reversible en determinadas ocasiones). Asimismo, los amiloplastos (contenedores de almidón) pueden transformarse en cloroplastos, hecho que explica el fenómeno por el cual las raíces adquieren tonos verdosos al estar en contacto con la luz solar.
Estructura y abundancia

En las plantas superiores, la forma que con mayor frecuencia presentan los cloroplastos es la de disco lenticular, aunque también existen algunos de aspecto ovoidal o esférico. Con respecto a su número, se puede decir que en torno a cuarenta y cincuenta cloroplastos coexisten, de media, en una célula de una hoja; y existen unos 500.000 cloroplastos por milímetro cuadrado de superficie foliar. No sucede lo mismo entre las algas, pues los cloroplastos de éstas no se encuentran tan determinados ni en número ni en forma. Por ejemplo, en el alga Spirogyra únicamente existen dos cloroplastos con forma de cinta en espiral, y en el alga Chlamydomonas, sólo hay uno de grandes dimensiones.
En el interior y delimitado por una membrana plastidial interna, se ubica una cámara que alberga un medio interno con un elevado número de componentes (ADN plastidial, circular y de doble hélice, plastorribosomas, enzimas e inclusiones de granos de almidón y las inclusiones lipídicas); es lo que se conoce por el nombre de estroma. Inmerso en el se encuentran una gran cantidad de sáculos denominados tilacoides, que contienen pigmentos fotosintéticos en su membrana tilacoidal (cuya cavidad interior se llama lumen o espacio tilacoidal). Los tilacoides pueden encontrarse repartidos por todo el estroma (tilacoides del estroma), o bien, pueden ser pequeños, tener forma discoidal y encontrarse apilados originando unos montones, denominados grana (tilacoides de grana). Es en la membrana de los grana donde se ubican los sistemas enzimáticos encargados de captar la energía luminosa, llevar a cabo el transporte de electrones y sintetizar ATP.
Función
La más importante función realizada por los cloroplastos es la fotosíntesis, proceso en la que la materia inorgánica es transformada en materia orgánica (fase oscura) empleando la energía bioquímica (ATP) obtenida por medio de la energía solar, a través de los pigmentos fotosintéticos y la cadena transportadora de electrones de los tilacoides (fase luminosa). Otras vías metabólicas de vital importancia que se realizan en el estroma, son la biosíntesis de proteínas y la replicación del ADN.FASE LUMINOSA O FOTO QUIMICA
La energía luminosa que absorbe la clorofila se transmite a los electrones externos de la molécula, los cuales escapan de la misma y producen una especie de corriente eléctrica en el interior del cloroplasto al incorporarse a la cadena de transporte de electrones. Esta energía puede ser empleada en la síntesis de ATP mediante la fotofosforilación, y en la síntesis de NADPH. Ambos compuestos son necesarios para la siguiente fase o Ciclo de Calvin, donde se sintetizarán los primeros azúcares que servirán para la producción de sacarosa y almidón. Los electrones que ceden las clorofilas son repuestos mediante la oxidación del H2O, proceso en el cual se genera el O2 que las plantas liberan a la atmósfera.
Existen dos variantes de fotofosforilación: acíclica y cíclica, según el tránsito que sigan los electrones a través de los fotosistemas. Las consecuencias de seguir un tipo u otro estriban principalmente en la producción o no de NADPH y en la liberación o no de O2.
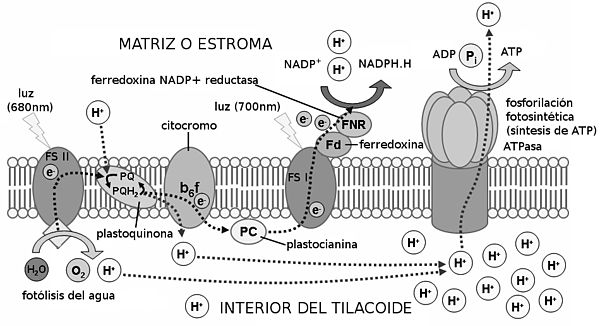
Los electrones pasan a una cadena de transporte de electrones, que invertirá su energía liberada en la síntesis de ATP. ¿Cómo? La teoría quimioosmótica nos lo explica de la siguiente manera: los electrones son cedidos a las plastoquinonas, las cuales captan también dos protones del estroma. Los electrones y los protones pasan al complejo de citocromos bf, que bombea los protones al interior del tilacoide. Se consigue así una gran concentración de protones en el tilacoide (entre éstos y los resultantes de la fotólisis del agua), que se compensa regresando al estroma a través de las proteínas ATP-sintasas, que invierten la energía del paso de los protones en sintetizar ATP. La síntesis de ATP en la fase fotoquímica se denomina fotofosforilación.
Los electrones de los citocromos pasan a la plastocianina, que los cede a su vez al fotosistema I. Con la energía de la luz, los electrones son de nuevo liberados y captados por el aceptor A0. De ahí pasan a través de una serie de filoquinonas hasta llegar a la ferredoxina. Ésta molécula los cede a la enzima NADP+-reductasa, que capta también dos protones del estroma. Con los dos protones y los dos electrones, reduce un NADP+ en NADPH + H+.
El balance final es: por cada molécula de agua (y por cada cuatro fotones) se forman media molécula de oxígeno, 1,3 moléculas de ATP, y un NADPH + H+.
El objetivo que tiene la fase cíclica tratada es el de subsanar el déficit de ATP obtenido en la fase acíclica para poder afrontar la fase oscura posterior.
Cuando se ilumina con luz de longitud de onda superior a 680 nm (lo que se llama rojo lejano) sólo se produce el proceso cíclico. Al incidir los fotones sobre el fotosistema I, la clorofila P700 libera los electrones que llegan a la ferredoxina, la cual los cede a un citocromo bf y éste a la plastoquinona (PQ), que capta dos protones y pasa a (PQH2). La plastoquinona reducida cede los dos electrones al citocromo bf, seguidamente a la plastocianina y de vuelta al fotosistema I. Este flujo de electrones produce una diferencia de potencial en el tilacoide que hace que entren protones al interior. Posteriormente saldrán al estroma por la ATP-sintetasa fosforilando ADP en ATP. De forma que únicamente se producirá ATP en esta fase.
Sirve para compensar el hecho de que en la fotofosforilación acíclica no se genera suficiente ATP para la fase oscura.
La fase luminosa cíclica puede producirse al mismo tiempo que la acíclica.
Existen dos variantes de fotofosforilación: acíclica y cíclica, según el tránsito que sigan los electrones a través de los fotosistemas. Las consecuencias de seguir un tipo u otro estriban principalmente en la producción o no de NADPH y en la liberación o no de O2.
Fotofosforilación acíclica
El proceso de la fase luminosa, supuesto para dos electrones, es el siguiente: Los fotones inciden sobre el fotosistema II, excitando y liberando dos electrones, que pasan al primer aceptor de electrones, la feofitina. Los electrones los repone el primer dador de electrones, el dador Z, con los electrones procedentes de la fotólisis del agua en el interior del tilacoide (la molécula de agua se divide en 2H+ + 2e- + 1/2O2). Los protones de la fotólisis se acumulan en el interior del tilacoide, y el oxígeno es liberado.Los electrones pasan a una cadena de transporte de electrones, que invertirá su energía liberada en la síntesis de ATP. ¿Cómo? La teoría quimioosmótica nos lo explica de la siguiente manera: los electrones son cedidos a las plastoquinonas, las cuales captan también dos protones del estroma. Los electrones y los protones pasan al complejo de citocromos bf, que bombea los protones al interior del tilacoide. Se consigue así una gran concentración de protones en el tilacoide (entre éstos y los resultantes de la fotólisis del agua), que se compensa regresando al estroma a través de las proteínas ATP-sintasas, que invierten la energía del paso de los protones en sintetizar ATP. La síntesis de ATP en la fase fotoquímica se denomina fotofosforilación.
Los electrones de los citocromos pasan a la plastocianina, que los cede a su vez al fotosistema I. Con la energía de la luz, los electrones son de nuevo liberados y captados por el aceptor A0. De ahí pasan a través de una serie de filoquinonas hasta llegar a la ferredoxina. Ésta molécula los cede a la enzima NADP+-reductasa, que capta también dos protones del estroma. Con los dos protones y los dos electrones, reduce un NADP+ en NADPH + H+.
El balance final es: por cada molécula de agua (y por cada cuatro fotones) se forman media molécula de oxígeno, 1,3 moléculas de ATP, y un NADPH + H+.

Fase luminosa cíclica
En la fase luminosa o fotoquímica cíclica interviene de forma exclusiva el fotosistema I, generándose un flujo o ciclo de electrones que en cada vuelta da lugar a síntesis de ATP. Al no intervenir el fotosistema II, no hay fotólisis del agua y, por ende, no se produce la reducción del NADP+ ni se desprende oxígeno. Únicamente se obtiene ATP.El objetivo que tiene la fase cíclica tratada es el de subsanar el déficit de ATP obtenido en la fase acíclica para poder afrontar la fase oscura posterior.
Cuando se ilumina con luz de longitud de onda superior a 680 nm (lo que se llama rojo lejano) sólo se produce el proceso cíclico. Al incidir los fotones sobre el fotosistema I, la clorofila P700 libera los electrones que llegan a la ferredoxina, la cual los cede a un citocromo bf y éste a la plastoquinona (PQ), que capta dos protones y pasa a (PQH2). La plastoquinona reducida cede los dos electrones al citocromo bf, seguidamente a la plastocianina y de vuelta al fotosistema I. Este flujo de electrones produce una diferencia de potencial en el tilacoide que hace que entren protones al interior. Posteriormente saldrán al estroma por la ATP-sintetasa fosforilando ADP en ATP. De forma que únicamente se producirá ATP en esta fase.
Sirve para compensar el hecho de que en la fotofosforilación acíclica no se genera suficiente ATP para la fase oscura.
La fase luminosa cíclica puede producirse al mismo tiempo que la acíclica.
FASE OSCURA O BIOSINTETICA

Síntesis de compuestos de carbono: descubierta por el bioquímico norteamericano Melvin Calvin, por lo que también se conoce con la denominación de Ciclo de Calvin, se produce mediante un proceso de carácter cíclico en el que se pueden distinguir varios pasos o fases.
En primer lugar se produce la fijación del dióxido de carbono. En el estroma del cloroplasto, el dióxido de carbono atmosférico se une a la pentosa ribulosa-1,5-bisfosfato, gracias a la enzima RuBisCO, y origina un compuesto inestable de seis carbonos, que se descompone en dos moléculas de ácido-3-fosfoglicérico. Se trata de moléculas constituidas por tres átomos de carbono, por lo que las plantas que siguen esta vía metabólica se llaman C3. Si bien, muchas especies vegetales tropicales que crecen en zonas desérticas, modifican el ciclo de tal manera que el primer producto fotosintético no es una molécula de tres átomos de carbono, sino de cuatro (un ácido dicarboxílico), constituyéndose un método alternativo denominado vía de la C4, al igual que este tipo de plantas.
Con posterioridad se produce la reducción del dióxido de carbono fijado. Por medio del consumo de ATP y del NADPH obtenidos en la fase luminosa, el ácido 3-fosfoglicérico se reduce a gliceraldehído 3-fosfato. Éste puede seguir dos vías, consistiendo la primera de ellas en regenerar la ribulosa 1-5-difosfato (la mayor parte del producto se invierte en esto) o bien, servir para realizar otro tipo de biosíntesis: el que se queda en el estroma del cloroplasto comienza la síntesis de aminoácidos, ácidos grasos y almidón. El que pasa al citosol origina la glucosa y la fructosa, que al combinarse generan la sacarosa (azúcar característico de la savia) mediante un proceso parecido a la glucólisis en sentido inverso.
La regeneración de la ribulosa-1,5-difosfato se lleva a cabo a partir del gliceraldehído 3-fosfato, por medio de un proceso complejo donde se suceden compuestos de cuatro, cinco y siete carbonos, semejante a ciclo de las pentosas fosfato en sentido inverso (en el ciclo de Calvin, por cada molécula de dióxido de carbono que se incorpora se requieren dos de NADPH y tres de ATP).
Síntesis de compuestos orgánicos nitrogenados: gracias al ATP y al NADPH obtenidos en la fase luminosa, se puede llevar a cabo la reducción de los iones nitrato que están disueltos en el suelo en tres etapas.
En un primer momento, los iones nitrato se reducen a iones nitrito por la enzima nitrato reductasa, requiriéndose el consumo de un NADPH. Más tarde, los nitritos se reducen a amoníaco gracias, nuevamente, a la enzima nitrato reductasa y volviéndose a gastar un NADPH. Finalmente, el amoníaco que se ha obtenido y que es nocivo para la planta, es captado con rapidez por el ácido α-cetoglutárico originándose el ácido glutámico (reacción catalizada por la enzima glutamato sintetasa), a partir del cual los átomos de nitrógeno pueden pasar en forma de grupo amino a otros cetoácidos y producir nuevos aminoácidos.
Sin embargo, algunas bacterias pertenecientes a lo géneros Azotobacter, Clostridium y Rhizobium y determinadas cianobacterias (Anabaena y Nostoc) tienen la capacidad de aprovechar el nitrógeno atmosférico, transformando las moléculas de este elemento químico en amoníaco mediante el proceso llamada fijación del nitrógeno. Es por ello por lo que estos organismos reciben el nombre de fijadores de nitrógeno.

- Síntesis de compuestos orgánicos con azufre: partiendo del NADPH y del ATP de la fase luminosa, el ión sulfato es reducido a ión sulfito, para finalmente volver a reducirse a sulfuro de hidrógeno. Este compuesto químico, cuando se combina con la acetilserina produce el aminoácido cisteína, pasando a formar parte de la materia orgánica celular.
FOTORESPIRACION

Cuando existe abundante dióxido de carbono, la enzima RuBisCO (mediante su actividad como carboxilasa) introduce el compuesto químico en el ciclo de Calvin con gran eficacia. Pero cuando la concentración de dióxido de carbono en la hoja es considerablemente inferior en comparación a la de oxígeno, la misma enzima es la encargada de catalizar la reacción de la RuBisCO con el oxígeno (mediante su actividad como oxigenasa), en lugar del dióxido de carbono. Esta reacción es considerada la primera fase del proceso fotorrespiratorio, en el que los glúcidos se oxidan a dióxido de carbono y agua en presencia de luz. Además, este proceso supone una pérdida energética notable al no generarse ni NADH ni ATP (principal rasgo que lo diferencia de la respiración mitocondrial).
Cuando una molécula de RuBisCO reacciona con una de oxígeno, se origina una molécula de ácido fosfogliceraldehido y otra de ácido fosfoglicólico, que prontamente se hidroliza a ácido glicólico. Este último sale de los cloroplastos para posteriormente introducirse en los peroxisomas (orgánulos que albergan enzimas oxidativos), lugar en el que vuelve a reaccionar con oxígeno para producir ácido glioxílico y peróxido de hidrógeno (la acción de la enzima catalasa catalizará la descomposición de este compuesto químico en oxígeno y agua). Sin embargo el ácido glioxílico se transforma en glicina, aminoácido que se traspasa a la mitocondrias para formarse una molécula de serina a partir de dos de ácido glioxílico (este proceso conlleva la liberación de una molécula de dióxido de carbono).
FOTOSISTEMAS Y PIGMENTOS FOTOSINTETICOS
Los fotosistemas
Los pigmentos fotosintéticos se hayan alojados en unas proteínas transmembranales que forman unos conjuntos denominados fotosistemas, en los que se distinguen dos unidas diferentes: la antena y el centro de reacción.
En la antena, que también puede aparecer nombrada como LHC, predominan las pigmentos fotosintéticos sobre las proteínas. De hecho, existen entre doscientas y cuatrocientas moléculas de pigmentos de antena de varios tipos y tan sólo dos proteínas intermembranales. Sin embargo, la antena carece de pigmento diana.
En el centro de reacción, mentado en algunas ocasiones como CC (abreviatura del inglés Core Complex), las proteínas predominan sobre los pigmentos. En el centro de reacción es donde está el pigmento diana, el primer aceptor de electrones y el primer dador de electrones. En término generales, se puede decir que existe una molécula de pigmento diana, unas cuantas de pigmentos no diana, una de primer dador de electrones y una de primer aceptor. Mientras existen entre dos y cuatro proteínas de membrana.
Los pigmentos fotosintéticos y la absorción de la luz
Los pigmentos fotosintéticos son lípidos que se hayan unidos a proteínas presentes en algunas membranas plasmáticas, y que se caracterizan por presentar alternancia de enlaces sencillos con enlaces dobles. Esto se relaciona con su capacidad de aprovechamiento de la luz para iniciar reacciones químicas, y con poseer color propio. En las plantas se encuentran las clorofilas y los carotenoides; en las cianobacterias y las algas rojas también existe ficocianina y ficoeritrina; y finalmente, en las bacterias fotosintéticas está la bacterioclorofila.
La clorofila está formada por un anillo porfirínico con un átomo de magnesio en el centro, asociado a un metanol y a un fitol (monoalcohol de compuesto de veinte carbonos). Como consecuencia, se conforma una molécula de carácter anfipático, en donde la porfirina actúa como polo hidrófilo y el fitol como polo lipófilo. Se distinguen dos variedades de clorofila: la clorofila a, que alberga un grupo metilo en el tercer carbono porfirínico y que absorbe luz de longitud de onda cercana a 630 nm, y la clorofila b, que contiene un grupo formilo y que absorbe a 660 nm.
Los carotenoides son isoprenoides y absorben luz de 440 nm, pudiendo ser de dos clases: los carotenos, que son de color rojo, y las xantófilas, derivados oxigenados de los nombrados anteriormente, que son de color amarillento. Las ficocianinas y las ficoeritrinas, de color azul y rojo respectivamente, son lípidos que se hayan asociados a proteínas originando las ficobiliproteínas.
Como los pigmentos fotosintéticos tienen enlaces covalentes sencillos que se alternan con enlaces covalentes dobles, se favorece la existencia de electrones libres que no pueden atribuirse a un átomo concreto.
Cuando incide un fotón sobre un electrón de un pigmento fotosintético de antena, el electrón capta la energía del fotón y asciende a posiciones más alejadas del núcleo atómico. En el supuesto caso de que el pigmento estuviese aislado, al descender al nivel inicial, la energía captada se liberaría en forma de calor o de radiación de mayor longitud de onda (fluorescencia). Sin embargo, al existir diversos tipos de pigmentos muy próximos, la energía de excitación captada por un determinado pigmento puede ser transferida a otro al que se induce el estado de excitación. Este fenómeno se produce gracias a un estado de resonancia entre la molécula dadora relajada y la aceptora. Para ello se necesita que el espectro de emisión del primero coincida, al menos en parte, con el de absorción del segundo. Los excitones se transfieren siempre hacia los pigmentos que absorben a mayor longitud de onda, continuando el proceso hasta alcanzar el pigmento fotosintético diana.
Suscribirse a:
Entradas (Atom)